3- الانسياب الكتلي :- Mass Flow
يتسسب الماء الخارج من الثغور في سحب تيار من الماء يبدأ من الثغر وينتهي إلى الجذر ثم محلول التربة وهذا التيار يتسبب في زيادة امتصاص الأيونات إما بصوره غير مباشرة عن طريق تحريك الأيونات بعد خروجها من أوعية الخشب مما يتسبب في زيادة نشاط الامتصاص الأيوني أو بصوره مباشرة بانسياب تيار الماء حاملا الأيونات من المحلول الأرضي خلال الجذور إلى الأفرع وقد تأيد تلك النظرية بتعريض الجذور لضغط هيدروستاتيكي فكانت النتيجه زيادة الأيونات الممتصة
4- تبادل الأيونات بالتلامس:- The contactExchange
فسر تلك النظرية القصور فى امتصاص الكاتيونات عن طريق نظرية CO2 فالامتصاص يتم أحيانا بدون وجود الكاتيون فى محلول التربة او عند pH مناسب ليحدث الامتصاص نتيجة لفرق الجهد ولكن تستطيع الجذور اخذ الكاتيونات مباشرة دون المرور بالمحلول الارضى فلكل أيون مدمص له حقل يتذبذب فية FieldOscillation فأذا تداخل مع مجال ذبذبة لأيون آخر مدمص على سطح غروى آخر او على أسطح الأغشية السيتوبلازمية للجذور فانه يحدث التبادل وبهذا يمكن ان تنتقل الأيونات مباشرا من حبيبة الطين الى الجذر حيث ان الجدار جدار السليولوز ليس خاملا حيث لارتباطه بمجموعة حامضية والجدار السليولوزى غير نقى لتخلل المسافات البينية له باللجنين والمواد البكتينية والتى لها خواص تبادلية
الانتقال النشيط :-
لوحظ أن امتصاص الأيونات يحدث بدون تدخل الطاقة الأيونيه إلى حد معين ثم يستمر بعد ذلك بصوره لا يمكن لنظريات الامتصاص السلبي تفسيرها وهذا الامتصاص التالي يحدث فيه تراكم للأيونات ضد تدرج التركيز ويتم تثبيطه عندما يكون النشاط الأيضي للنبات مثبطا بالحرارة المنخفضة أو تركيز الأكسجين المنخفض أو المثبطات الأيضيه ….إلخ . وهنا يمكن افتراض أن التراكم الأيوني في النبات يحتاج إلى طاقه أيضيه وانتقال الأيونات بالأستعانه بالطاقة التمثيلية ويرمز له باصطلاح الامتصاص النشيط Active Transportion عن طريق حوامل Carrier موجودة في الأغشية الخلوية .
مـا هـو الحـامـل Carrier Concept ؟
أعتقد البعض أن هناك ممرات خلال المنطقة بين الحيز الخارجي والداخلي للخلية لا تكون منفذه للأيونات الحرة تحتوي على حوامل منخفضة هذه الحوامل ترتبط مع الأيونات في الحيز الخارجي ثم تتخلص منها في الحيز الداخلي وأكثر ملامح نظرية الحوامل أهميه هو افتراض وجود معقد الحامل والأيون وهو مركب يمكنه التحرك خلال الغشاء الغير منفذ السابق ذكره وعند تتحرك الأيونات ولا تستطيع الحامل الحركة مره أخرى للخارج وعلى ذلك يتم تراكما بينما يعود الحامل فارغا . وهناك ثلاث خصائص تؤيد بشده صدق مفهوم الحوامل .
1- تبادل النظـائر :- Isotopic Exchange
وجد أن الأيونات الممتصة امتصاصا نشيطا تكون غير قابله للتبادل مع الأيونات من نفس النوع في الحيز الخارجي أو الوسط الخارجي وقد لوحظ ذلك بأستخدام النظائر المشعة مما يدل على أن الغشاء ذو مناعة مرتفعه للأتنشار الحر للأيونات وحيث أن الأيونات تمتص فإن حركتها خلال الغشاء يجب أن تمر بواسطة وسيط هو الحوامل .
وقد تم دراسة امتصاص الكبريت المعلم كب 35 بواسطة جذور الشعير ووجد أن الكميه الممتصة يمكن أن تفصل إلى جزئـين :-
1- كمية تمتص بالأنتشـــــــــار
2- كمية تمتص بالامتصاص النشط
كما وجد أن الكميه الممتصة بالأنتشار يمكن أن تخرج عند غمر النسيج في الماء خروجا حرا بينما الكميه الممتصة امتصاصا نشيطا لا تخرج إلى الماء كما لا يمكنها التبادل مع أيونات كبريت عاديه في محلول كبريت غير مشبع
الامتصاص النشط ........ز الانتشار بالمساعدة ......... الامتصاص النشط ............انتشار خلايا الاغشية الدهنية
شكل يوضح انواع الامتصاص حيث يمتص النبات مغذياتة بطريقتين اما امتصاصا سلبيا او امتصاصا نشطا
الامتصاص تقف عند حد معين أو عند نقطة التشبع عندها تكون جميع المواضع النشطة على الحوامل محتله ، وهي حالة متشابه لتأثير التشبع المعروف في التفاعلات الأنزيمية ، وهناك حقيقة تقول ان مستوى المعدل الأقصى للامتصاص يمكن أن يستمر لفترة طويلة نسبيا يؤيد وجود عدد محدود من الحوامل العاملة بأقصى كفاءه بمعنى أن المواضع النشيطة على الحوامل تكون مشغولة طول الوقت . وبمجرد أن يتخلص أحد الحوامل من الأيون في الحيز الداخلي فإنه في الحال يتم انشغاله بأيون آخر من منطقة الحيز الخارجي للنسيج وعلى ذلك فإنه عند نقطة التشبع تظل الدورة مستمرة ولا يمكن أن تعمل بسرعة أكبر لو أن تركيز الأملاح تزايد .
3 - الـتــخصـص :- Specificity
الجذور تمتص الأيونات اختياريا بمعنى أن معدل امتصاص الأيونات يختلف كما يختلف معدل تراكمها في أنسجة الجذور مما يؤيد وجود الحوامل المتخصصةSpecificity Carriers هذه الخاصية ترتبط بالأيونات ذات السلوك الكيماوي الغير متشابه إلا أنها لا تظهر مع الأيونات المتشابه أو تكون ضعيفة الأثر فمثــلا الكاتيونات الأحادية مثل البوتاسيوم والسيزيوم والروبيديوم تتنافس مع بعضها على نفس موضع الالتحام على الحامل أي أن معدل امتصاص الروبيديوم يمكن أن يخفض بإضافة مزيد من البوتاسيوم أو السيزيوم إلى المحلول المغذي وزيادة تركيز الروبيد يوم يمكن أن يتلاشى الأثر المثبط للكاتيونات الأخرى ومما يؤيد وجود مواضع التحام مختلفة أن وجود زيادة من الصوديوم لا يثبط امتصاص الروبيديوم ، كذلك فإن السلينات تثبط امتصاص الكبريتات بينما لا تؤثر على امتصاص الفوسفات أو النترات . وهي حاله تتشابه أيضا مع علاقة الأنزيم السيسترات حيث يفسر على أساس انتقال المواضع الفعالة في الأنزيم .

ميكـانيكيـة الامتصاص النشـيط
هناك ميكانيكيتان محتملتان لعمل الحوامل :-
1- مضـخـة السيتوكروم : Cytochrome Pump
لوحظ Hoagland واخرين أن غمر النسيج في محلول محلي يتسبب في زيادة معدل التنفس وهو ما يسمى بالتنفس الملحي وان امتصاص الاملاح يعتمد على التنفس وهي ما قادLundagardh 1950 إلى نظرية تصيغ العلاقة السابقة ومحتواها هو ان الامتصاص يحدث من خلال السيتوكروم اوكسيديز هذا بالاضافة الى ان السيتوكروم قد يكون حاملا للأيونات وقد تأكد من ذلك عندما وجد ان الامتصاص يثبط بتثبيط التنفس بمثبطات السيتوكروم اوكسيديز فالحديد بالسيتوكروم يغير تكافؤه من ثنائى الى ثلاثى والعكس فأذا كان ثلاثى ح +++ يأخذ الكترون او انيون سالب ليصبح ح ++ والعكس وان مصدر الأيدروجين هو الأحماض العضوية الموجودة بجدار الاندوبلاست تحت تأثير أنزيم Dehydrogenase
وعلية فان :
1- امتصاص الأنيونات يعتبر مستقلا عن امتصاص الكاتيونات وكلاهما يحدث عن طريق ميكانيكيات مختلفة
2- يوجد تدرج في تركيز الأكسجين من السطح الخارجي الى السطح الداخلي للغشاء منها عند السطح الداخلـي .
3- الانتقال الفعلي للانيونات يحدث من خلال نظام السيتوكروم.
وحيث أن هناك علاقة كميه بين امتصاص الأيونات والتنفس الملحي وهذه العلاقة لا تظهر عند امتصاص الكاتيونات فقد أفترض أن أيونات فقط تنقل انتقالا نشيطا . وتثبط التنفس الملحي وما يترتب عليه من تثبيط امتصاص الأملاح بواسطة أول أكسيد الكربون قادت لاند جارد لان يفترض أن انتقال الأيونات يتم بواسطة السيتوكروم اوكسيديز وبالتالي فإن السيتوكروم قد يكون من الحوامل الأنيونية .

شكل يوضح الية الامتصاص تبعا لنظرية الحوامل او نظرية مضخة السيتوكروم التى تفترض وجود حامل ينشطة ATP يغير من شكله فينفتح على الخلية من الداخل ليمتص منها الصوديوم ثم يغير من شكله ليفرز الصوديوم خارجها ويمتص بدلا منه البوتاسيوم او اى كاتيون آخر ثم يغير شكله مرة آخرى ليتمكن من افراز الكاتيونات داخل
يتبع.................







 رد مع اقتباس
رد مع اقتباس


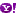





 ......................
......................

مواقع النشر (المفضلة)